Vorbemerkung
Eduard Jacobshagen (* 12. Mai 1886 in Hämelschenburg; † 22. Dezember 1968 in Marburg) war ein deutscher Anatom und Professor an der Universität Marburg.
Von 1906 bis 1911 studierte Jacobshagen Naturwissenschaften und Medizin an den Universitäten Jena und München. Von 1915 bis 1918 diente er im Ersten Weltkrieg. 1912 promovierte er, 1919 habilitierte er sich für Anatomie, Ver-gleichende Anatomie und Entwicklungsgeschichte an der Universität Jena. 1921 wechselte der Privatdozent nach Leipzig und wurde dort 1924 ao. Professor für Anatomie. Seit 1926 hatte er einen Lehrstuhl für Anatomie in Marburg. Er befasste sich mit der Anatomie der Wirbeltiere und mit Merkmalen der nordischen und friesischen „Rasse“. 1945 wurde er entlassen, aber 1949 wieder eingestellt und 1957 erneut emeritiert.
Seit Mai 1933 war er Mitglied der NSDAP, daneben im NS-Ärztebund, im NS-Lehrerbund, in der NSV. Im November 1933 unterzeichnete er das Bekenntnis der deutschen Professoren zu Adolf Hitler. Vor und nach dem Krieg engagierte er sich in der Bodendenkmalpflege.
(Wikipedia, Stand 15. April 2021)
Es ist möglich, daß in seiner NSLB-Mitgliedschaft auch die Verbindungen zu Luttropp in Ziegenhain und Boley in Fritzlar ihre Ursache haben. Ersterem verdankt die Forschung die Entdeckung des Quarzit-basierten Mittel-paläolithikums in Nordhessen schon in den 1930er Jahren, was sicher auch Erwartungen genährt haben mag. Auch Boley reagiert in "Das Kasseler Becken und seine Randgebiete in vorgeschichtlicher Zeit" schon 1960/61 recht bald auf den Rhündaer Fund (s. Regionalmuseum-Fritzlar-virtuell/Veröffentlichungen).

Unwetter richteten am 19./20. Juli 1956 im Einzugsgebiete des Rhünder Baches schwere Schäden an. Dieser Bach mündet gegenüber der Schloßruine Altenburg in den Unterlauf der Eder unmittelbar unterhalb der Schwalmmündung. Tosende Wasserfluten hatten sich während der Nacht etwas oberhalb des Fundplatzes ein neues Bett geschaffen. Am rechten Rande dieses neuen Bachlaufes war 75-80 cm unter der Erdoberfläche offenkundig die linke Hälfte eines auf seiner rechten Seite liegenden und hier mit Kalkstein außen sehr versinterten menschlichen Oberschädels mitgerissen. Herr Glatzer, Hauptlehrer in Rhünda, kam am frühen Morgen des 20. Juli, um die Hauptschäden seines Dorfes mit Hilfe seiner Kamera festzuhalten. Da wurde er vom Ortseinwohner Schober auf den vom Wasser noch etwas überspülten Schädelrest aufmerksam gemacht. Der Schädelrest und die Fundschichtoberfläche wurden zunächst in dieser Situation photographiert. Darauf wurde das Fundstück von Herrn Glatzer mit dem Taschenmesser umstochen, vorsichtig geborgen und ins Schulhaus gebracht. Herr Glatzer erreichte vom Bürgermeister alsbald die Absperrung des Fundplatzes, bis dessen wissenschaftliche Untersuchung erledigt war. So ist erfreulich sachgemäß gehandelt worden. Am 22. Juli früh fuhr Herr Glatzer in Urlaub. Bereits am 22. Juli nachmittags erschienen auf eine irrtümliche Zeitungsmitteilung hin die Herren L. Köhler und Fr. Rödde von der Musealen Arbeitsgemeinschaft für Ur- und Vorgeschichte des nahen Fritzlar in Rhünda. Diese beiden überaus einsatzfreudigen Herren fuhren nach Besichtigung des Fundplatzes unter der Mitnahme von Fundschichtproben, des Fundstückes und der Mutter des Herrn Glatzer mit ihrem Kraftwagen zu mir nach Marburg in die Wannkopfstraße weiter. 2 Tage später war der Fund soweit trocken, daß ich ihn bearbeiten konnte.
Zu meiner Person: Seit 43 Jahren bin ich Anatom, seit 34 Jahren auch Anthropologe. Urmenschenkunde war hier mein Hauptarbeitsgebiet. 1928 nahm ich die Erforschung der nordhessischen eiszeitlichen Tierwelt auf, und suchte durch Lehrausflüge zu meiner Vorlesung über Eiszeitmenschen, teils in nahes Gelände, teils in berühmte alte Fundstätten, nachhaltigeres Sachinteresse zu wecken. 1941 nahm ich die planmäßige Erforschung des gesamten Naturgeschehens einer Eiszeit in Nordhessen auf. Ein glänzender Fundplatz und ein starker Arbeitseinsatz hatten mein -Mühen 1950 zu einem vollen Erfolge gelangen lassen. Alsbald begann ich, Nordhessens Eiszeitkunde in seine Heimatkunde mit Eiszeit-Ausstellungen und mit an 100 Lichtbildvorträgen und mit Führungen an den bedeutsamsten meiner Fundplätze einzubauen. So gelang es in steigendem Maße, Mitarbeit im Lande wachzurufen. Ihr dankt die Wissenschaft auch den vorliegenden Fund. Ich habe dann die großen Zusammenhänge im eiszeitlichen Naturgeschehen des Erdballs aufdecken können. Dies benutzte ich, Nordhessens eiszeitliche Heimatkunde in ihren weltweiten Rahmen zu stellen, als Empiriker Wissenschaft zu lehren, „die dem Menschen nützt, indem sie ihn erhebt“ (Goethe). Es war kosmisches Geschehen übersehbar geworden! Ich durfte erleben, daß Adolf Luttropp, Lehrer in Ziegenhain, ab 1938 die Aufdeckung der eiszeitlichen Menschenkulturen hier in einer für Deutschland einzigartigen Weise gelang, und daß er sich nach 15jährigem Ringen zunächst international durchzusetzen vermochte. Der Unterstützung durch den Heimatbund für Kurhessen und Waldeck verdanken wir es, daß wir beiden unsere Erkenntnisse in fünf gemeinsam veranstalteten Eiszeiten-Ausstellungen allein an mehr als 20000 Menschen haben herantragen können. - Seit 15 Jahren wollte ich ferner zeigen, daß dem Mangel an eiszeitlichen Fundstücken aus deutschem Boden in erster Linie durch rühriges Sammeln abgeholfen werden kann und muß. Nach vielen Tausenden zählen Luttropps eiszeitlichen Menschengeräte wie meine eiszeitlichen Tierfunde heute! Darum beschritten wir neue Wege, weil der rasch fortschreitende rnaschinelle Abbau auf den deutschen Fundplätzen unseres Erachtens gar keine andere Wahl ließ. Wir drängten zu besseren Einsichten.
Der Fundplatz des Schädels liegt nordwestlich des durch seine Basaltbrüche bekannten Dorfes Rhünda, südöstlich der Bahnstrecke Marburg-Kassel. Genauer: Halbwegs zwischen der Landstraße Harle-Gensungen und der Bahnstrecke Wabern-Gensungen. Wer den Fundplatz aufsuchen und die Bahn benutzen will, steigt von Richtung Kassel her in Gensungen-Felsberg aus. Kommt er aus Richtung Marburg oder aus dem Waldeckischen, so ist Wabern die Aussteigestation. Mit Kraftwagen fährt man von der Bundesstraße Kassel-Marburg ab Fritzlar über Wabern-Harle; vom Fiildatale her entweder von Beiseföri,h westwärts über lihünda, oder von-Melsungen her über Gensungen links zur Kreuzung mit, der Rhündaer Straße. Den gleichen Weg nimmt man von der Autobahn Kassel-Frankfurt.
Erdgeschichtliches: Die Besichtigung des neuen Laufes des Rhünder Baches am Fundplatz lehrt, daß das neue Bett in eine alte Talaue eingeschnitten ist. Denn in den Seitenwänden wie im Boden stecken abgerollte Basaltblöcke sehr verschiedener und oft bedeutender Größe (über 40 cm). Zwischen ihnen mehr oder weniger grobsandiges Material. Ich habe diese Talwand als Aufschüttung einer glazialen Tundrenzeit im Sinne W. SOERGELS aufgefaßt. Über ihr Alter wage ich kein Urteil, denn wir befinden uns kaum 200 m vom Rande der noch bewegten niederhessischen Senke.
Eine 40-50 cm starke Lößlehmschicht mit Schottern heimischer Gesteine bildet das Hangende der Fundschicht. Sie dehnt sich auch über die nordostwärts unmittelbar angrenzenden Äcker bis hin zur Eder. Südwärts der Straße Harle-Gensungen liegt sie, laut der Geologischen Karte, Blatt Gudensberg, zwischen Harle und der Gensunger Zementfabrik, und ist von Basaltschottern meist durchsetzt. Hier kommen neben Basaltdurchbrüchen verschiedener mineralogischer Zusammensetzung westlich Rhünda Inseln oligocäner Ablagerungen, östlich Rhündas solche miocäner Tone und Sande vor, stellenweise mit Braunkohlenquarziten.
Die Fundschicht ist deutlich gegen den hangenden Lößlehm und ebenso nach der Tiefe zu abgesetzt. Ihr Liegendes zeigt oft eine grob-wellige Oberfläche, die mir mit alten Erdflußerscheinungen zusammenzuhängen scheint. Der Fundschicht ist eigentümlich, daß sie unzusammenhängender Verbreitung hat. Diese merkwürdige Erscheinung meinte ich mit Abtragungsvorgängen in Zusammenhang bringen zu dürfen. Trocknet man Fundschichtproben, so sehen sie stumpftorffarben aus und haben einen humosen Einschlag. Ihre physikalische Beschaffenheit ist vielfach eine krümelige, an anderen Stellen zeigt sie gröbere und grobe, durch Kalksinter zusammengeballte Klumpen. Auch die krümelige Beschaffenheit geht wohl auf Versinterung zurück. Die Hauptmasse der Kalksteinniederschläge geschah um Busch- oder Baumwurzeln, die inzwischen vergangen sind, und deren eines Ende dann manchmal zusinterte. Neben Wurzelkanälen unter 0,5 mm Durchmesser maß ich solche bis zu 4 cm und fand auch grobe Wurzelverzweigungen. Vielfach ziehen sich um größere Achsenkanäle mehrere konzentrische Sinterlamellen. Zwischen sie sind manchmal etwas Sand oder kleine Konkretionsbröckchen geschaltet. Außen sind wohl auch Quarz und kleine Basaltbrocken in sie eingebacken. Aus durchweichten Klumpen holte ich mehrere Zentimeter lange, stark abgerollte, Basaltstücke heraus. Aus härteren wurden sie durch Anschleifen nachgewiesen. Die eingesinterten Steine haben meist eine eigene dünne Kalksteindecke. Aus einem weicheren Sinterklumpen, den ich mit einem starken Pfriem auseinanderbrach, kam das versinterte Gehäuse einer Radix ovata Draltarnaud ans Licht.
Als altem Fossiliensammler aus Lößlehm war mir bekannt, daß Kalksteinkonkretionen sich stets zuerst auf der Unterseite von Gegenständen niederschlagen, die sich dem Wege kalkhaltigen Wassers entgegenstellen. Meine hernach zu schildernde Erfahrung in der Fundschicht lehrte, daß das kalkhaltige Wasser von oben und mit einer Tendenz bachabwärts bewegt worden ist. Es hat also den Weg des Niederschlagswassers genommen.
Könnte es aus dem jetzt überlagernden Lößlehm gekommen sein?
Nein! Dann wäre ja die Oberfläche der Fundschicht nicht klar und deutlich eben gewesen. Dann hätte ich niemals eine Radix ovata aus den Sintern holen können. Also mußte die Sinterbildung vor sich gegangen sein, ehe der Löß der Deckschicht sich niederschlug. Denn das aride Klima einer Tundrenzeit schloß eine Sinterbildung aus! Nur in einem wärmeren Klima dominiert chemische Bodenverwitterung und -auslaugung in der hier vorauszusetzenden Form. Also konnte die Fundschicht ihren Charakter nicht erst im Alluvium erhalten haben, sondern muß eine interglaziale oder interstadiale Entstehung besitzen! Der Fundschicht ist, laut Angabe des Herrn Glatzer, der Schädelrest 25-30 cm unter der Oberfläche entnommen, ungefähr gleich hoch über ihrem Liegenden. Auf Grund der Versinterurgserscheinungen an ihm, muß der Schädel in seiner Fundschicht versintert sein. Ob er sich hier in sekundärer Lagerung befindet oder in primärer, lasse ich offen.
Woher kommt der Kalkgehalt, der Fundschicht?
Der Rhünderbach führt heute kein hartes Wasser. Für ein spätes oder mittleres Pleistocän wird man angesichts der geologischen Zeitnähe in der Hinsieht keinen anderen Zustand voraussetzen wollen. Der große Kalkreichtum der Fundschicht hält, mich ebenso davon ab, an Auswaschungen aus einer sehr alten Lößbedeckung zu glauben, die vor der Abtragung oberer Teile der auf alle Fälle in einem wärmeren Klima gebildeten Fundschicht bestanden hätte, doch sonst restlos verschwunden wäre. Da jedoch unter dem Lößlehm der Gegend vielfach tertiäre Gesteinsschichten anstehen, wird man meines Erachtens ohne weiteres annehmen dürfen, daß Kalkansschwemmungen aus Melanienton vom Hange des südöstlich liegenden Rhünder Berges hier zur Auskristallisation von Kalkstein gezwungen wurden.
Noch ist die Frage zu streifen, welcher Eiszeit der Lößlehm entstammt? Man wird meines Erachtens an ein Produkt mehrerer Eiszeiten denken dürfen, welches vom Gegenwartsklima in den heutigen Zustand versetzt wurde. Nichts nötigt wohl, die Schotter des Lößlehms ins gleiche Glazial zu stellen wie den Lößniederschlag.
Der Fund ist gut 5 m über dem Ederspiegel geborgen worden. So wird man annehmen dürfen, daß die Fundschicht einer Fußterrasse aufliegt.
Deshalb habe ich das geologische Institut der Universität Marburg gebeten, die genaue erdgeschichtliche Fundplatzbearbeitung zu übernehmen, was mir zugesagt worden ist.
Der Zustand des Fundstückes:
Abb. 1 zeigt die rechte Seite des Rhündaer Schädels nach seiner Bergung. Das Oberkiefer- und Nasengebiet ruht in der rechten Hand des Jungen. Seine linke liegt über der Hinterhälfte des linken Scheitelbeins, Die Knochenstücke oberhalb des Ringbruches im Schädeldache sind hier nicht vorhanden. Die neben dem rechten Daumennagel des Jungen aufsteigende Sinterfurche lag unterhalb des Jochbogens. Der helle Wulst, rechts davon, bedeckte den Jochbogen und die Unterhälfte der Fossa temporalis. Davon rechts (im Schatten) die Oberhälfte der Ala magna ossis sphenoidis. Der Rundbuckel über dem rückwärtigen Jochbogenende lagerte über dem Oberende der Pars mastoidea des Schläfenbeins und dein anschließenden Unterende des Scheitelbeins.
Das Photo, das Herr Glatzer von dem auf seiner rechten Seite liegenden Fundstück in situ gemacht hat, zeigt mir, daß zunächst alles so an seinem Platze zu finden war, wie es nun nach meiner Instandsetzung zu sehen ist. Isoliert mir übergebene Schädeldachstücke sind erst beim Bergen oder beim Trocknen aus ihrer Verbindung herausgefallen.
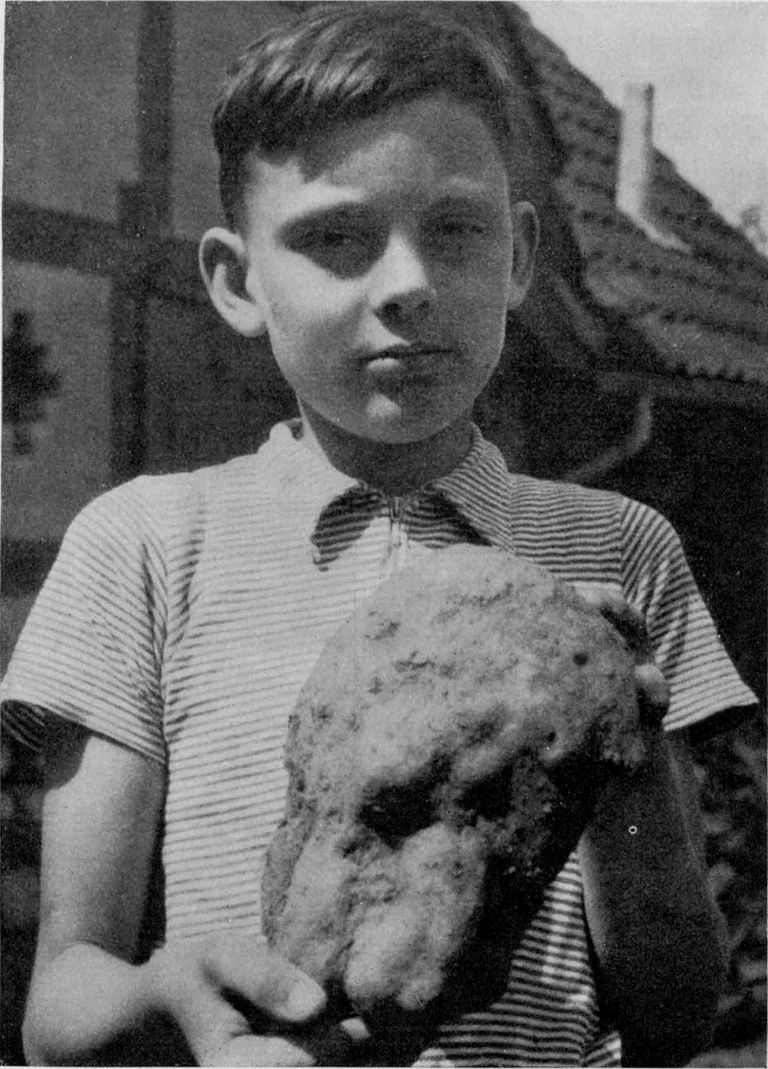
Abb. 1. Die rechte Seite des
Fundstückes in der Sinter-
hülle.
(Photo Fröhlich)
Da waren zunächst senkrecht durchschneidende Sprünge im Schädeldach (Abb. 5). Sie gingen von einem Punkte links des Bregma aus. Sie haben den Zusammenhalt inr Kuppelbau der Scheitelbeine schwer gefährdet. Dies um so mehr, als das Schädeldach außerdem einen schiefliegenden Ringbruch aufwies. Er begann in der Median-Sagittalen in der oberen Hälfte des Scheitelbeins, zog knapp fingerbreit unter dein rechtien Foramen parietale, dann dicht über dem Oberrande der Schläfenschuppe hinweg nach vorn und schräg durch die Squama frontalis nach links. Der schiefsitzende Ringbruch, wie die radiäre Zerklüftung des Schädeldaches, können nur durch eine stauchende Gewalteinwirkung auf das bereits versinterte Schädeldach entstanden sein. Innerhalb des Ringbruches fehlte bei der Fundbergung die sonst dem Schädel fest anhaftende Sinterdecke! Dieser begegnete ich erst unterhalb des Ringbruches. Vielleicht war es in der Unwetternacht ein von rechts hinten anrollender Stein, der die Sprünge setzte und so das Abgeschlepptwerden des größten Teils der linken Craniumseite durch die Wasserströmung auslöste. - Linkerseits (Abb. 8) war das Hinterhauptsbein im wesentlichen erhalten. Vom Os parietale war nur der hintere untere Teil und das mediale Stück noch da, vom Schläfenbein dessen Pars mastoidea, während von deren Pars petrosa ein vorderer lateraler Teil zerstört war. Die Schläfenbeinschuppe, die linke Keilbeinhälfte, die linke Maxilla, beide Nasenbeine, das linke Jochbein und das linke Stirnbein fehlten gänzlich. Im vorderen Parietalbereiche griffen die Defekte auf die rechte Seite über, ebenso im hinteren medialen Teile der rechten Stirnbeinschuppe sowie im Dache und im Boden der rechten Orbita. Der Schädel drehte sein Hinterhaupt dein Dorfe zu, das Gesicht, bachabwärts.
Als ich die Bearbeitung des Fundstückes begann, machte ich erst an einem kleinen Knochensplitter einen Entkalkungsversuch mit 10% Salzsäure, um zu sehen, ob das Fundstück fossil sei. Danach begann die Meißelarbeit. Der Schädel war fossil. Er zeigte sich dann beim Meißeln als recht hart und spröde. Vor dem Meißeln wurde das Fundstück auf einen Haufen dünner Leintücher gelegt, so daß es eine gewisse Verschieblichkeit erhielt. Mit scharfen Meißeln zog ich dann erst radiäre flache Rillen, die ich dann langsam vertiefte, ehe ich mit horizontaler Meißelstellung an die Abtragung der Sintermassen heranging. Betrug am Ringbruch die Dicke der Sinterschicht 5-6 mm, so wuchs ihre Stärke einerseits in Richtung auf den Jochbogen, d. h. in Richtung auf den tiefst gelegenen Schädelteil, aber stärker noch nach der ederwärts gewandten Gebiß- und Gaumenseite, wo die Mächtigkeit der Bedeckung durch Sinter 30 mm erreichte. So fand sich die Massierung der Versinterung so verteilt, wie sie erwartet werden mußte, wenn Regen von oben her in die Fundschicht gedrungen und dann auch bergab gesickert war. Zwischen Jochbein und, Schläfengrube hatte sich mir eine seichte Furche angedeutet. Sie bewog mich, zunächst von oben-hinten her den Jochbogen freizumeißeln. Leider platzte dabei die Warzenfortsatzgegend ab (Abb. 5). Sie war linksseitig vorhanden. Deshalb habe ich den technisch schwierigen Versuch unterlassen, die Sinterhülle vom rechten Processus mastoides zu entfernen. Im vorderen oberen Quadranten der Sprung-fläche hob sich das Antrum mastoideum durch einen feinen Sinterüberzug mit eingebackenen humosen Sanden deutlich ab. Dieses Sintermaterial ist offenbar von der Fossa cranii media her durch eine Reihe feiner Öffnungen lateral der Eminentia arcuata an seinen Ort gelangt. Eine der dortigen feinen Öffnungen fand ich durch Sinter zugesetzt. - Beim Ausmeißeln der Sinterdecke im Bereiche der Fossa canina (Abb. 6) sprang leider deren dünner Boden heraus. Ich habe dann von innen her mit Gips eine stärkere Außenwand der Kieferhöhle geschaffen, auch im Bereiche der Facies infratemporalis sowie in der Gegend der Incisura lacrimalis. Defekte im Bereiche des Keilbeins und seiner Oberkieferverbindung wurden durch Gips und durch Leimungen verstärkt und ausgeglichen. Nach Freilegung der Austrittsstellen im Bereiche der mittleren Schädelgrube habe ich zur Verstärkung des Ganzen deren Boden im Verlaufe des Sulcus n. petrosi sup. majoris eine Gipsauflage gemacht. Bemerkt sei noch, daß der Boden der Fossa cranii occipitalis bis über den Sulcus transversus hinaus, und ebenso der Boden der mittleren Schädelgrube mit einer dünnen Sinterdecke gleichmäßig bedeckt war (im Raume der F. er. occipitalis auch linkerseits!).
Imprägniert habe ich mit Zell-Auto-Klarlack mit entsprechender Verdünnung. Wo Uhu sich beim Kleben als unzu-länglich erwies, ist Celloluidpulver mit Azeton angewandt worden.
Allgemeine erste Angaben über das Cranium: Die größte Schädellänge betrug 197 mm. Bei der Messung wurden wegen eines Knochendefektes der Tabula externa an der Glabella 2 mm zugerechnet. Die tiefgelegene größte Breite maß 140 mm. Wir haben also einen Langschädel mit dem Längen-Breitenindex 71 vor uns (Ultra-dolichocephal).
Das Lebensalter dürfte zwischen 35 und 40 Jahren gelegen haben. Die Bregmagegend fehlt. Die allein vorhandenen rückwärtigen Abschnitte der Sutura sagittalis waren vollkommen verstrichen. Die Pars temporalis der Koronarnaht war offen, ebenso die Sutura sphenofrontalis. An der Lambdanaht war die Pars asterica im Schließen.
Das Geschlecht muß das weibliche gewesen sein. Die Schädelknochen waren dünn: nur 4 mm an der Stirnbeinschuppe hinter dem Torus supraorbitalis, 4,5 mm vor dem Hinterrand des Os parietale links. Dazu war das Muskelrelief besonders in der Nackenregion wenig kräftig. Auch die bescheidene Länge der rechten Reihe der fünf Molariformes mit 40 mm wird man so deuten müssen. Zuletzt erinnert die Norma verticalis stark an kindliche Verhältnisse, wie das beim weiblichen Geschlecht oft gefunden wird.
Zur Anatomie der Schädelknochen.
1. Os occipitale.
a) außen: Pars basilaris: Sie ist, 2,6 cm lang, vorn links beschädigt, mit deutlichem Tuberculum pharyngicum (Abb. 2).
Partes laterales: Condylus 1. fast zerstört, r. vorn etwas verletzt. Größter Transversaldurchmesser 23 mm, in Sagittalrich- tung kurz. Er springt 14,5 mm in das Foramen magnum vor! Letzteres ist vorn zugespitzt. Ein wohl einmaliger Befund.
Squama: Protuberantia und Linea nuchalis terminalis sind schwach (Abb. 5), die Linea supraterminalis ist nur angedeutet.
b) innen: Pars basilaris: Clivus flach längs eingemuldet.
Partes laterales: Mit Tuberculum jugulare.
Sqnama: R. Sulcus transversus steht mit seinem Oberrande etwas höher als die Linea nuchalis terminalis.
2. Os parietale.
Vorn rechts fehlt dem Knochen ein stumpf dreieckiges Stück (Abb. 7). Seine Länge beträgt am Medialrande knapp 2,5 cm. Einen leicht S-förmig geschwungener Rand kehrte es nach lateral-hinten. In der Kranznaht fehlten etwas über 20 mm.

Abb. 2. Rhünda. Vorderteil
der Schädelbasis. 1/2 natür-
liche Größe.
a) außen: Kein Parietalhöcker. Linea temporalis sehr schwach. Foramen parietale nur rechts.
b) innen: Prächtige Sulci arteriosi (Abb. 8). Geringe Foveolae granulares hinter dein Foramen parietale. Sulcus sagittalis flach (Abb. 6).
3. Os temporale:
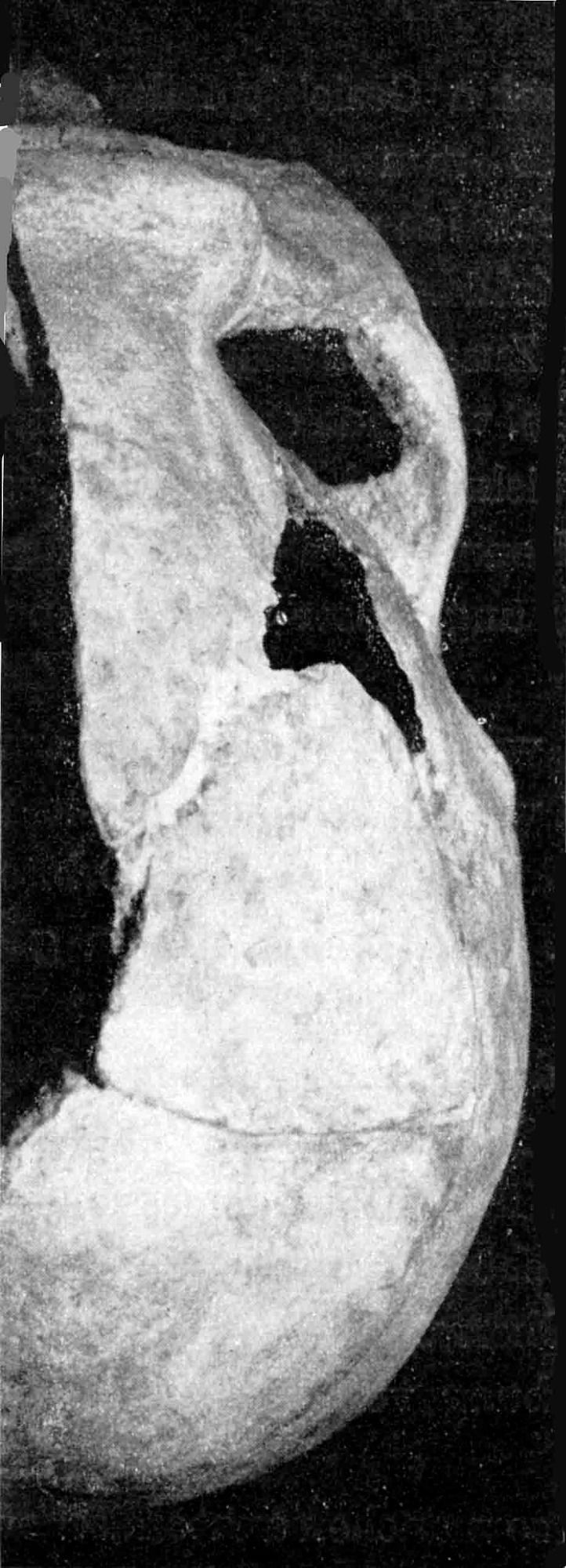
Vorn oben fehlt etwas vom Rande der Squama (Abb. 5). Der rechte Warzenfortsatz ist abgesprungen, der linke aber vorhanden (Abb. 8). Dort, fehlt aber fast die gesamte Squama.
a) außen: Squama: Eine Rinne für die Arteria temporalis media fehlt. Processus zygomaticus durch Sutur mit Jochbein ver- bunden. Auf seiner Unterseite das Tuberculum articulare und die 8,5 mm tiefe Fossa mandibularis. Sie steht mit ihrem Grunde 5 mm unter dem Porion (Abb. 5).
Pars mastoidea: Sutura squamoso-mastoidea fehlt. Länge des Warzenfortsatzes, medial gemessen, 15 mm. Er ist pneumatisiert, Antrum mastoideum wohl entwickelt (Abb. 3). Foramen mastoideum.
Pars tympanica: Links fehlt der vordere obere Rand. Rechts nur ein Rest derselben, die untere Hälfte ist vorhanden. Meatus acusticus läuft nach medial und senkt sich in der inneren Hälfte, zumal hinten, links mit Kalksteinkonkretionen (Abb. 3).
Pars petrosa: Unterfläche: Fossa jugularis tief, halb geschieden vom Foramen caroticum externum. Versinterungen in beiden (Abb. 2). - Processus styloidei beiderseits kurz abgebrochen. Foramen stylomastoideunm links frei. Incisura mastoidea vorhanden, oberhalb Sulcus der Arteria occipitalis.
b) innen: Squama mit schönen Sulci arteriae meningicae med. (Abb. 8). Pars petrosa: Rück- und Vorderfläche o. B.
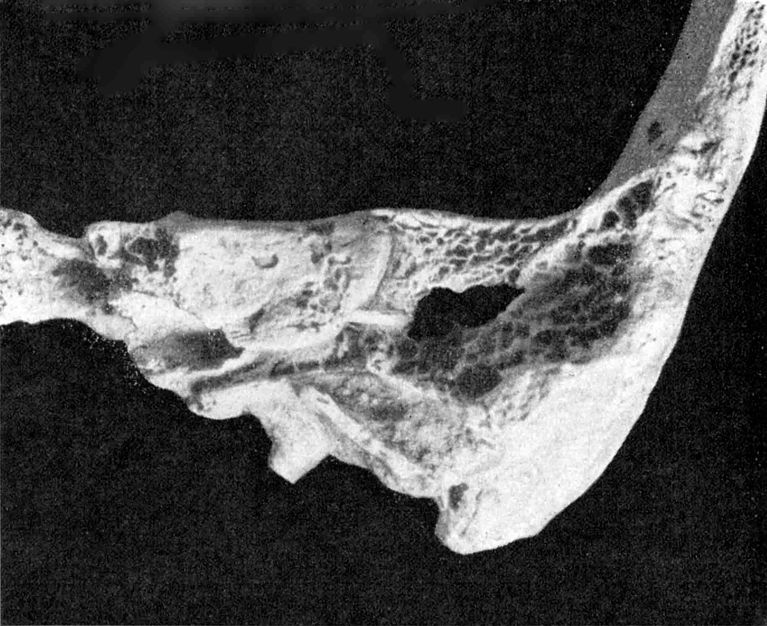
Abb. 3. Rhünda. Linksseitige
Bruchfläche durch das knö-
cherne Ohr.
Auf der linken Schädelseite gibt ein Schrägbruch eine ganz vorzügliche Übersicht über die knöchernen Räume des Ohres.
Abb. 3 zeigt am Grunde des äußeren Gehörganges deutlich den Sulcus für die Trommelfellanheftung. Letzteres stand nach außen-unten und etwas nach vorn geneigt. Medialwärts von ihm blicken wir in das niedrig-trommelförmige Cavum tympani mit dem Promontorium auf seiner Medialwand. Nach miedial-vorn gehen Semicanalis musculi tensoris tympani und darunter der Semicanalis tubae pharyngotympanicae vom Cavum aus. Lateral-hinten wird das Cavum vom lateralen Bogengang überwölbt, und über seinem Vorderende findet sich die Eminentia arcuata mit dem von lateral her sichtbarem oberen Bogengang. Hinter und lateral des Canalis semicircularis lateralis sieht man das Antrum mastoideum. Seine Tiefe stellte ich mit 2 cm fest. Auch die Antrumverbindung mit dem Cavum tympani liegt vor dem Beschauer. Aus der Nähe erkennt dieser auch den Hiatus nervi facialis.
4. Os frontale.
Rechts ist die Glabella gerade noch da (Abb. 6). Rückwärts von ihr fehlt ein Knochenstück in Form eines stumpfwinkligen Dreiecks. Seine längste Seite dreht es der Medianebene zu, während die kürzeste an das Os parietale grenzt. Das fehlend, Stück hat bis 16 mm Breite.
a) außen: Den Oberrand der Augenhöhle nimmt ein lateral wohl entwickelter Torus supra-orbitalis ein, den nach medial- oben eine sanfte Furche von einem knapp 2 cm langen Arcus superciliaris abzugrenzen scheint. Letzterer erreicht die Glabella, auf der reichlich 1,5 mm der Tabula externa abgestoßen sind. Die Squama ist sehr stark geneigt (Abb. 8), die aus dem lateralen Torusende sich entwickelnde Linea temporalis kräftig.
b) innen: In der Fossa cranii frontalis hinten schöne Sulci arteriosi des vorderen Astes der A. meningica media. Vorn die Sulci der durch die Lamina cribriformis eingetretenen Arteria meningica anterior aus der A. ethmoidalis anterior in gleicher Schönheit. Kein Sinuns frontalis (Abb. 8).
5. Os sphenoides.
a) außen: Großenteils im Bild der Norma lateralis zu ersehen (Abb. 5). Die Nähte nach vorn, oben und hinten waren offen. Unten, seitlich von der Ala magna ausgehend, ein 1 cm langes Basisstück der beiden Flügelfortsätze! Die Fossa zwischen ihnen war ehedem von kleinen Steinchen erfüllt, die miteinander versintert waren.
b) innen: Es fehlen: Dorsum sellae, Fossa hyhophyseos, Tuberculum sellae, Sulcus und Canalis fasciculi optici, ebenso der kleine Keilbeinflügel (Abb. 8). Erhalten blieb ein Stück der rechten Seitenwand des Knochens mit Teilen des Sulcus caroticus, die Ala magna mit Canalis rotundus, Foramen ovale und Foramen spinae. Infolge der Beschädigung des Keilbeinkörpers liegt dessen Hohlraumsystem frei. Die rechte Keilbeinhöhle erscheint zweigeteilt. Die vordere dieser Höhlen gehört deutlich zur vorderen Keilbeinanlage, deren kleiner Flügel an unserem Fundstück fehlt. Die doppelt so große hintere Keilbein- höhle gehört ebenso deutlich der hinteren Keilbeinanlage zu. Der Vorderwand des Keilbeinkörpers fehlt das Rostrum. Ein Lateralstück der Concha und die Apertura sinus sphenoidei ist jedoch erhalten.
Im Bereiche des Gesichtsschädels fehlen: Os ethmoidale, Os nasale, Os lacrimale, Os masillotiurbinale und Vomer.
1. Os maxillare.
a) außen: Das Corpus führt einen großen Sinus. Auf dem Tuber liegen drei Foramina alveolaria. Der Processus frontalis ist über der Apertura piriformis abgebrochen. Fossa pränasalis, die Spina nasalis fehlt (Abb. 6). Der Processus zygomaticus o. B. Der Processus palatinus hinten mit Sulci versehen. Processus alveolaris in parabolischem Verlaufe (Abb. 2). Die vordere und die mediale Wand der Alveole des J 1 fehlt. 6 mm dahinter findet sich der Rinnenanteil des Canalis incisivus. Kleine Konkretionen an der Lateralwand der Alveole des J 1 zeigen, daß der J 1 vor der Fossilisierung fehlte. In der Alveole des J 2 stecken versinterte kleine Steinchen, ebenso in der Alveole des Caninus, deren Vorderwand beschädigt ist.
P 1, P 2, M 1, M 2 und M 3 sitzen in den Alveolen. Am M 1 und M 2 sind die beiden bukkalen Wurzeln deutlich vonein- ander getrennt (Abb. 5).
Die Angaben über die einzelnen Zähne mögen hier gleich folgen. Die Kauflächenlänge der fünf Molariformes beträgt 40 mm. Die größte Länge der einzelnen Kauflächen ist: P 1 = 4,5 mm, P 2 = 5 mm, M 1 = 7,7 mm, M 2 = 9,6 mm, M 3 = 9,0 mm. Die Kronenhöhen: P 1 = 2,2 mm, P 2 = 2,8 min, M 1 = 3,1 mm, M 2 = 5,2 mm, M 3 = 5,0 mm. Die größte labiolinguale Kronenbreite mißt: P 1 = 8,1 mm, P 2 = 10,2 mm, M 1 = 12,9 mm, M 2 = 12,4 mm, M 3 = 11,8 mm. Die Kaufläche des M 3 steht ziemlich horizontal, die der übrigen Molariformes steigen palatinal an, am meisten M 1 (Abb. 2 und 5).
b) innen: Auf der Medialfläche des Oberkiefers ist der Vorderrand des Hiatus sinus maxillaris ziemlich erhalten. Sein oberer und hinterer Rand fehlt. Die Medialfläche des Processus frontalis ließ ich unter ihrer Sinterdecke stehen, um die Bruchgefahr des Ganzen herabzusetzen.
2. Os palatinum.
a) außen: Die Lamina palatina ist nur in ihrer lateralen Hälfte erhalten. Dort ist das Forarnen palatinum majus sichtbar (Abb. 2).
b) innen: Es fanden sich nur untere Reststücke der Pars perpendicularis. Sie wurden zur Festigung des Ganzen eingegipst.
3. Os zygomaticum.
Es ist wohl entwickelt und durch seinen kräftigen Processus frontosphenoides bemerkenswert. Hinter ihm, wie hinter seinem Processus temporalis und dessen rückwärtiger Gelenkverbindung, ließ ich einen Teil der Sintermasse zur Festigung des Ganzen stehen.
Ein Blick in die Orbita.
Lateral: Außer der Facies orbitalis des Jochbeines ist auch die des großen Keilbeinflügels erhalten. Auf letzterer ließ ich wegen ihrer Zartheit die Sinterbedeckung stehen.
Oben : Die Facies orbitalis des Stirnbeins ist nahezu vollständig. Medial:Os ethmoidale und Os lacrimale fehlen.
Unten: Die Facies orbitalis maxillae sprang mir beim Meißeln heraus. Ich wünschte den Sinus maxillaris offenzuhalten und setzte den Knochen nicht wieder ein.
Zur Anthropologie des Fundstückes.
Nach unserem heutigen Wissen waren alle dem Homo sapiens LINNÉ vorausgehenden Menschenarten durch Platycephalie ausgezeichnet. Für das Schädeldach des Homo neandertalensis KING hat das ja besonders GUSTAV SCHWALBE klargelegt (1901). Seine Abbildung der Mediankurve der Neandertalkalotte mit vorn spitzem Bregma- und hinten spitzem Lambdawinkel über der Glabella-Inion-Linie (zur Protuberantia) und sein Kalottenhöhenindex verbinden sich uns Anthropologen zuerst mit dem Begriffe der Plattschädeligkeit der Neandertalmenschen.
Durch die Platycephalie wird die gesamte Schädel Form eindrucksvoll abgewandelt (Abb. 5.). Das Cranium sieht aus, als sei es von seinem Dache her gegen seine Basis gedrückt. Es übertrifft die größte mittlere Länge, die heute bei Homo sapiens gefunden wird. Der Betrag dieser zusätzlichen Länge liegt für Männer in Europa - La Chapelle, Monte Circeo, Spy 1, Neandertal, Spy II, Krapina und Le Moustier - zwischen 21 und 9 mm, für Frauen - La Quina, Ehringsdorf, Rhünda und Gibraltar - zwischen 24 und 11 mm. Für javanische Neandertaler liegt er - Ngandong V, XI, I und VI - zwischen 34 und 5 mm; für den afrikanischen Schädel von Broken Hill bei 23 mm. Am Cranium von Rhünda liegt die Schädellänge mit 197 mm um 18 mm über dem höchsten Mittelwert, der weiblichen Schädellage des heutigen Menschen.
Die Stirnneigung des Rhündaer Schädels ist sehr groß, wenngleich sie in beiden Normae laterales infolge des Fehlens eines schmalen medialen Stirnbeinstreifens etwas zu flach erscheint (Abb. 5 und 8).
Die Wölbung des Planum occipitale ist bei der Kalotte aus dem Neandertal sehr gering. Der Wölbungsgipfel (das Opisthocranium) liegt der Protuberantia occipitalis externa ganz nahe, nämlich auf 1/10 der flöhe des Lambda über der Glabella-Inion-Linie. Am Cranium der Frau von Rhünda ist das Planum occipitale stärker gewölbt und das Opisthocranium liegt auf 4/10 der Lambdahöhe. Die Hinterhauptsabplattung am Rhündaer Schädel ist auch im unteren hinteren Scheitelbeinbereiche etwas geringer als bei der Neandertalkalotte. Damit, fällt unser Fundstück jedoch keineswegs aus dem Formenkreise der Neandertaler heraus. Hier ist zunächst hervorzuheben, daß die Krümmungsverhältnisse am Hinterkopf der Rhündaerin sich ähnlich verhalten wie am Cranium des 8jährigen Kindes von La Quina und beim 16jährigen Jüngling von Le Moustier (vgl. WEINERT 1932, Abb. 76). Womit diese Formeigentümlichkeit der- Rhündaerin zusammenhängt, wird beim Studium der Großhirnverhältnisse gezeigt werden. Daß das Schädeldach des etwa 5jährigen Neandertaler Kindes aus Gibraltar im Hinterhaupt - wie im Stirnbereich jedoch wesentlich steiler als bei unserem Fundstück aufsteigt, sei erinnert.
Der Begriff der Platycephalie umfaßt heute längst weit mehr als die Zustände am Schädeldach! Studieren wir seine Fortentwicklung! RUDOLF VIRCHOWs „Untersuchungen über die Entwicklung des Schädelgrundes“ des Homo sapiens von 1857 hatten den Grund des Schädels, den Träger des Gehirns, als seinen formbeständigsten Teil erkannt. Nur eine wichtige Umgestaltung wies er an ihm auf, eine postnatale Basisknickung. Sie vollziehe sich bis zum Anbruch der Pubertät und falle vorzüglich in die zwei ersten Lebensjahre. HERMANN WELCKER bestimmte das mittlere Maß dieser Basisknickung im männlichen Geschlecht mit 7o.
Vergleichend-anatomisches Denken hat viel später diese Entdeckungen der Aufhellung des Werdeganges der menschlichen Schädelform dienstbar zu machen verstanden. Auf THOMAS HENRY HUXLEY, der in seinen Aufsätzen von 1863 „Zeugnisse für die Stellung des Menschen in der Natur“ als erster die Sonderstellung des Neandertalers klar übersehen hat, geht der fruchtbringende Gedanke über die Platycephalie zurück. Keine Schädelvergleichung sei viel wert, „die nicht auf die Bestimmung einer verhältnismäßig fixierten Grundlinie“ zurückgeführt werde. Diese liege in der relativen Konstanz der Schädelbasisverhältnisse vor 1). Aus Medianschnitten von Schädeln, die er plazentalen Säugetieren entnommen hatte, ging ihm auf, daß sich die Längsachse der Tier-Schädelbasis im Verhältnis zur Großhirnlänge verkürze, wenn er sie nach ihrer tieferen oder höheren systematischen Stellung betrachtete. Er führte dies am Beispiel eines Schädels vorn Biber, eines Lemuren und eines Pavians vor Augen. Andere Autoren prüften die Richtigkeit dieser Lehre nach. Sie brachten die stärkere Basisknickung einerseits mit einer sich steigernden Hirnentfaltung, andererseits mit der Erwerbung des aufrechten Ganges in Verbindung. So schrieb RUDOLF MARTIN 1928 im zweiten Bande seines Lehrbuchs der Anthropologie: „Eine stärkere basale Knickung geht immer mit einer stärkeren Gehirnentwicklung Hand in Hand, und auch die durch die Aufrichtung bedingte Kopfstellung der Hominidan hat ihren Einfluß auf die Schädelhasis ausgeübt.“ - Wir werden sehen, daß die erste der MARTINschen Behauptungen an entscheidender Stelle sich als irrig erweist: bei Homo sapiens! Daß auch die zweite in dieser Verallgemeinerung nicht gültig ist, haben WEIDENREICH und 1931 DABELOWs Untersuchungen klargestellt. ARTHUR KEITH unterließ das Spekulieren nach den Ursachen der Erscheinungen. Er ging statt dessen in seiner „Antiquity of Man“ (1925) den vorkommenden Konstruktionen und den dynamischen Zusammenhängen an den Cranien des Homo sapiens und des Homo neandertalensis mit vollem Erfolge nach. Auf sie muß ich hier näher eingehen.
Abb. 4 gibt KEITHs anschaulich erläuternde Schemareichnungen der Zustände bei Homo sapiens und bei Homo neandertalensis wieder, da mir entsprechende aus dem deutschen Schrifttum unbekannt sind. Punktiert ist das Schädelbasisverhalten eingetragen.
Gehen wir vom Schädelbasisbild des Neandertalers aus! SERA hat den Keilbeinwinkel des Gibraltarschädels - ihn nimmt KEITHs Abbildung zum Muster – mit 140° bestimmt. Das entspricht den Maßen, die WELKER beim Neugeborenen des Homo sapiens gefunden hat 2). Die Neandertaler behalten also zeitlebens die Basisknickung bei, die das
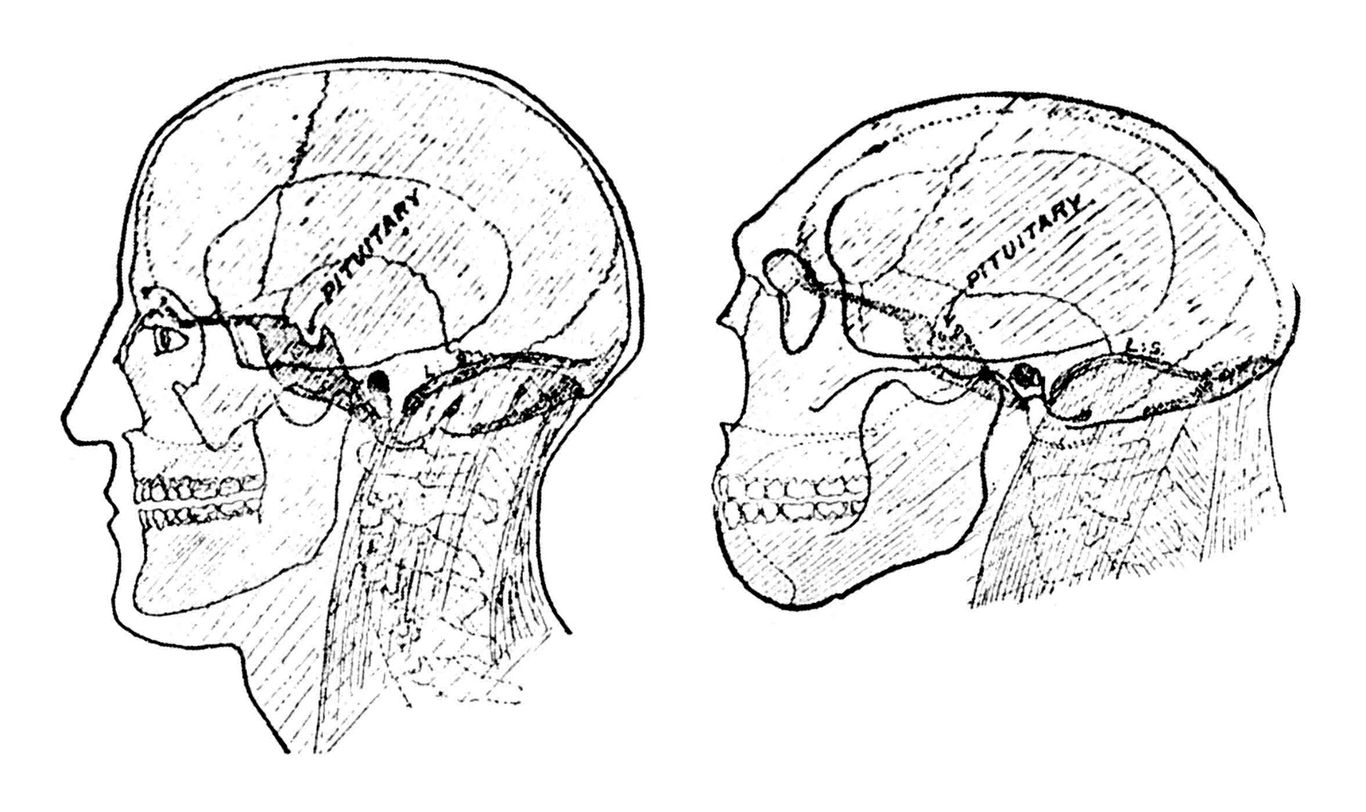
Abb. 4. Schädel von Homo sapiens und Homo neandertaliensis, lialbdurchsdig gezeichnet
nach A. KEITH 1925.
Neugeborene des Homo sapiens aufweist, die aber dann einer stärkeren Basisknickung weicht. Da die erwachsenen Neandertaler eine größere Schädelkapazität aufweisen als die erwachsenen Homo sapiens, ist die stärkere Basisknickung bei Homo sapiens also in keiner Weise an eine Größenzunahme des Großhirns gebunden, wie man nach -MARTIN glauben soll.
Angesichts der technischen Schwierigkeiten, die einer zuverlässigen Bestiminung des Keilbeinwinkels bisweilen begegnen, ist es erfreulich, daß bei Anwesenheit eines stumpferen Keilbeinwinkels der als Fossa mandibularis bezeichnete Teil der Pfanne des Unterkiefergelenkes eine tiefere Stellung zum Porus acusticus einnimmt, als beim spitzeren Keilbeinwinkel des erwachsenen Homo sapiens. Steht der Boden der Fossa mandibularis beim Neandertaler mit seiner typischen Platycephalie des Schädeldaches und mit seinem stumpfen Keilbeinwinkel nahe der Gehörgangsmitte, so bei Homo sapiens nahe dein Oberrande des Gehörganges und bei den Anthropopoiden nahe dessen Unterrande. Diese meines Erachtens höchst wichtigen Angaben A. KEITHs habe ich seit 30 Jahren überall, wo es mir möglich war, nachgeprüft! Sie stimmen für Homo sapiens. Ich kann auch die Angaben bestätigen für folgende Neandertaler - Männer: Spy I, Spy II, Le Moustier und La Chapelle; Frauen: Gibraltar, Ehringsdorf und Rhünda (Abb. 5).
Wir werfen nun den Blick auf den Kieferapparat! (Abb. 4). Beim Neandertaler ragt er vorn schnauzenartig vor. Sein die Zähne tragendes Corpus mandibulae ist breit und kräftig. Der Ramus mandibulae hat sich unter dein Einfluß einer entwickelteren Kaumuskulatur breiter und derber entwickelt. Der Angulus mandibulae biegt aus gleichem Anlaß steiler empor. Condylus und Processus muscularis sind gleichfalls wuchtiger geworden als bei Homo sapiens.
Wir denken uns nun aus dem Neandertalerschädel den Unterkiefer herausgenommen und sein Cranium aus Knetgummi geformt! Seinen Keilbeinwinkel wollen wir im Geiste um 7° spitzer machen. Da haben wir denn das steile Sapienshinterhaupt und die steile Sapiensstirn aus der Neandertaler-Platycephalie hervorgezaubert! Nicht stärkere Entfaltung des Großhirns gestattete das. Der Kauapparat wurde schwächer! Solange der Condylus mandibulae eines Homo neandertalensis KING in die Fossa mandibularis glitt, war sein Keilbeinwinkel keineswegs zu dem eines Homo sapiens LINNÉ zu machen! Indem wir uns die hier waltende Korrelation klarmachen, weiten sich die Platycephalie der Neandertaler, die des Homo heidelbergensis - ich erinnere hier an den geistvollen Versuch JOHANNES SOBOTTAs -, die des sogenannten Pithecanthropus und die des sogenannten Sinanthropus pekinensis zum Ausdruck der umklammernden Grundform des Gesamtschädels der Vorwohner des Homo sapiens LINNÉ auf unserem Planeten! Zugleich erkennen wir einen fundamentalen Einfluß des Gebißgebrauchs auf die Formgestalt der menschlichen Schädelbasis.
Wir aber verbleiben nun im platycephalen Formenkreis des Homo neandertalensis KING, wie dies unser neuer einzuordnender Schädelfund erfordert. Seine Norma lateralis (Abb. 5) - das ist hier von großer Bedeutung - zeigt uns die Fossa mandibularis 5 mm Unterhalb des Porion, wie sich das für einen Neandertaler gehört! Das ist darum erfreulich, weil der Keilbeinwinkel infolge von Beschädigungen an unserem Fundstück nicht exakt zu bestimmen ist (vgl. Abb. 8).
Unabziehbar vom Begriff des Neandertalmenschen ist seit langem der ihn kennzeichnende Torus supraorbitalis. Der Forschung ist es gelungen, auch ihn als Attribut der Platycephalie zu begreifen. Hier spielte C. TOLDTs Arbeit, von 1914 eine Hauptrolle.
In der Norma lateralis des Schädels von Rhünda zieht das laterale Torusende, nach Neandertaler Art, in den Processus zygomaticus der Stirnbeinschuppe hinein. Leicht kolbig angeschwollen biegt es dort nicht nur ab-, sondern auch rückwärts und überwölbt die Sutura spheno-frontalis (vgl. auch Abb. 6). Wir blicken auf den charakteristischen Pufferungswulst des Kaudrucks der drei Oberkiefermolaren, der über den Processus zygomaticus maxillae mit Hilfe des sehr kräftigen Processus fronto-sphenoides des Jochbeins von unten und schräg-rückwärts gemeinsam der Linea temporalis und - größeren Teils - dem Torus zugeleitet wird. Dabei sichert der Jochbogen diesem „jugalen Gesichtspfeiler“ in seiner Vertikalstellung.
Die lange fliehende Stirn des Neandertalers anstelle der mehr oder minder steil gestellten Squarna frontalis des Homo sapiens, schließt eine unmittelbare Kaudruckübertragung in das federnde Hirnschädeldach weitgehend aus. Der Oberrand der Orbita verstärkt sich „unabhängig von der Anlage der Stirnhöhlen“ in seiner Tabula externa. Hier entstünde eine fein-
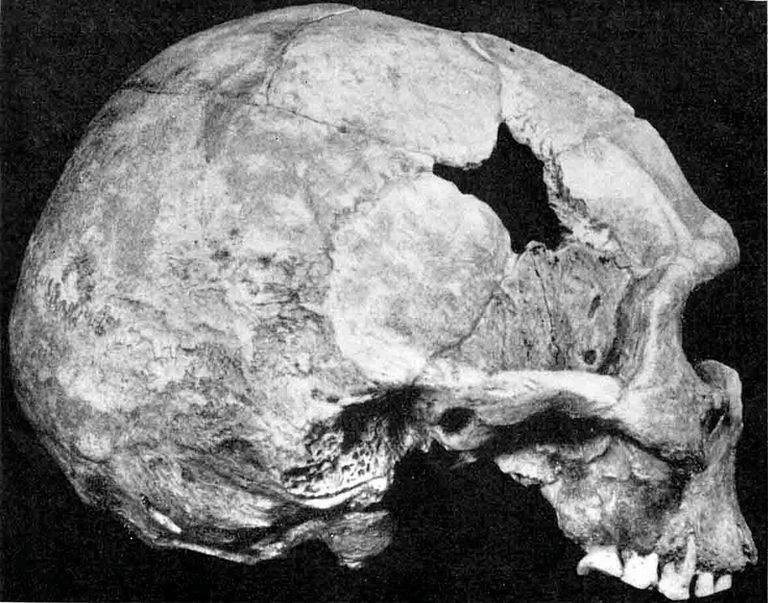
Abb. 5. Rhünda Norma late-
ralis r. 1/2 natürliche Größe
(4 mmm zu kurz).
blättrige, lockere, dem spongiösen Gewebe ähnliche Knochensubstanz mit einer besonderen, vom Periost herkommenden Blutversorgung. Im vorgerückteren Alter erst könne sich hier auch kompakter Knochen bilden. Später zeigt das Röntgenbild auch Knochenbögen, denen die Druckübertragung auf das flachstehende Stirnbein obliegt (O. GÖRKE 1904). Der Grad der Rückwärtsneigung der Stirn wie die Intensität der Gebißarbeit bestimmen den Entwicklungsgrad des Torus der Neandertaler.
Fiel in der Norma lateralis mancher Neandertalerschädel eine Spitzpoligkeit des Os occipitale in Höhe der Protuberantia externa bei einem flachen Hinterhaupt unter dem Inion auf, wie in der Regel bei Neugeborenen des Homo sapiens (Broken Hill), so lädt das Planum nuchale bei anderen (La Chapelle) rückwärts aus wie bei Sapienskindern des ersten Lebensjahres. Bei dritten tat dies auch (las Planum occipitale wie bei Sapienskindern zwischen dem 2.-5. Jahre. Von der Fixierung eines Entwicklungsstadiums der Bildung der Hirnschädelform des Homo sapiens in der entgültigen Schädelform der Neandertaltmenschen kann keine Rede sein. Die große Schädellänge entstand auch bei ihm erst im Pubertätsalter. Den Neandertalerschädel beherrscht in erster Linie eins, die Platycephalie, die in der typischen Basisknickung ersten Ausdruck findet.
Die Stirnansicht des Fundstückes verlangt einige erläuternde Hinweise. Die Glabella ist erhalten. Der Processus frontalis der Maxilla ist beim Kleben mit dem Oberende zu weit zur linken Kopfseite verschoben. Eine Beschädigung hat nicht nur die Vorderwand der Alveolen des J 1, J 2 und des C erlitten, sondern der J 1-Alveole fehlt auch die Medialwand. Eine Tangente an den spongiösen Knochen, gleich unterhalb der Apertura piriformis wie an den freien Glabellarand gelegt,
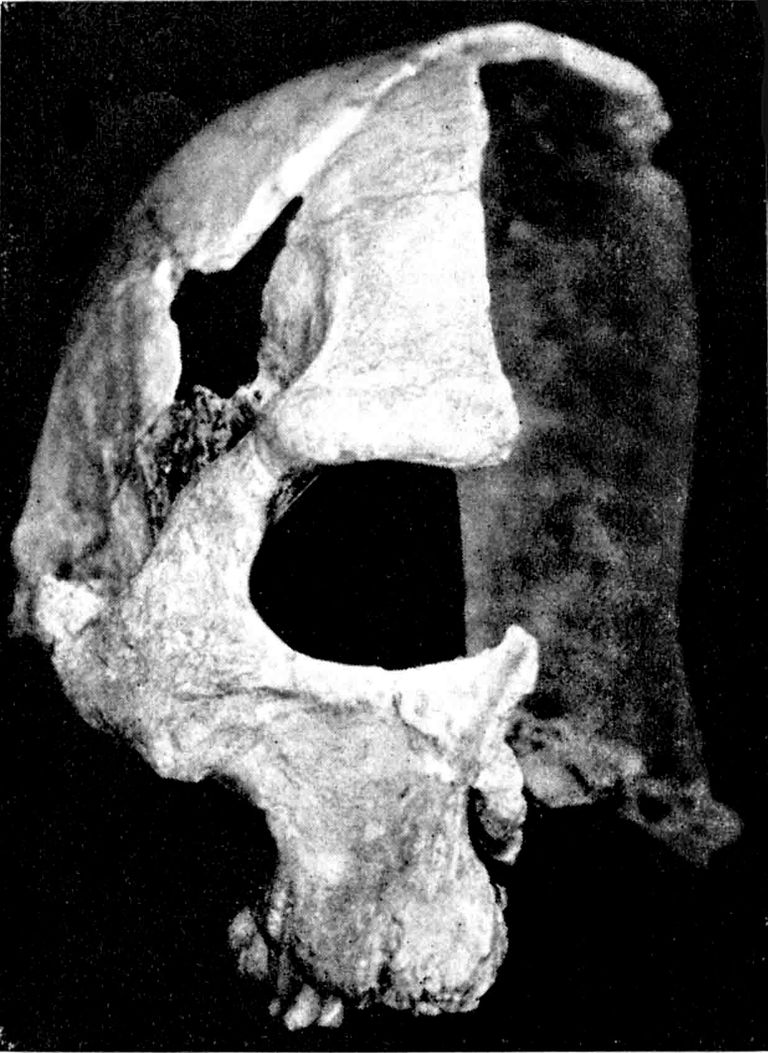
Abb. 6. Rhünda, Norma fron-
talis, 1/2 natürliche Größe.
trifft rechtsseitig nahe der Bruchspalte im Os parietale den Grund des Sulcus sagittalis, und damit die Mediansagitale des Schädeldaches.
Am typisch gebauten, von rechts nach links durchlaufenden Torus supraorbitalis bezeichnet eine schwache Schrägfurche nahe seinem medialen Drittel den Außenrand des 2 cm langen Arcus superciliaris. Von seiner kolbigen lateralen Anschwellung war schon die Rede. Die Torusbreite erreicht dort 1 cm. Das ist für erwachsene Neandertaler ein durchaus bescheidenes Maß.
Ein von einer Jochbeinnaht zur andern kontinuierlich durchlaufender Torus supraorbitalis, der keine Incisura frontalis lateralis oder höchstens eine Andeutung von ihr zurückließ, ist mir von folgenden Neandertalern bekannt. Männer: Le Moustier, Neandertal, Spy I, Spy II, Krapina, La Chapelle und Broken Hill; Frauen: Gibraltar, La Quina, Ehringsdorf und Rhünda.
Vorzüglich tritt der „nasale Gesichtspfeiler“ heraus. Er leitete den Beißdruck der Incisivi und des Caninus sowie den Kaudruck der Prämolaren einst über den Stirnfortsatz der Maxilla, medial der Orbita, auf den durch den Torus verstärkten Stirnbeinunterrand fort. Schon im stabilen Unterrande der Orbita verband er sich zu brillenartiger Umfassung der Augenhöhle mit dem „jugalen Gesichtspfeiler“, und diese Stabilisierung des unpaaren mit den paarigen Pfeilern erhielt im glabellaren Torusabschnitt, wie bei allen Neandertalerschädel, ihre letzte Sicherung gegen Querbruch 3).
Deutlich liegt zwischen dem jugalen und dem nasalen Gesichtspfeiler eine flachgemuldete Fossa canina. Sie soll beim Neandertaler immer fehlen. Dieses vermag ich zurzeit nicht nachzuprüfen. Hervorheben möchte ich aber die bei Homo sapiens so unterschiedlichen Befunde wegen dieser Mulde. Ihren angeblichen Mangel bei den Neandertalern in einen ursächlichen Zusammenhang mit der „schnauzenförmigen Bildung des Oberkiefers“ zu bringen, wie -man es bei MARTIN liest, ist angesichts der Fossa canina bei Negern undiskutabel. Gut wird es sein, das „Fehlen“ der Fossa canina nicht voreilig als Kennzeichen der Neandertaler zu deklarieren! Was wissen wir von der Bedeutung dieser Grube denn Sicheres?
Auf die geringe Stirnbreite unseres Fundstückes wird bei Erörterung der Norma verticalis eingegangen. Klar erkennt man, daß wir ein schmales Gesicht vor uns haben. Dem entspricht der Abstand zwischen dem Unterrand der Orbita und dem Alveolarrand des Oberkiefers über der Mitte des P 2 mit 45 mm gemessen, durchaus.
Der Eingang der rechteckigen Orbita überrascht durch seine geringe lichte Höhe von 29 mm. Denn das sind 10 mm weniger als am Schädel von La Chapelle! Die orbitale Breite - vom Ektoconchion zum Maxillofrontale gemessen - mißt 36 mm. Das sind 11,5 mm weniger als beim Schädel von La Chapelle! Auch die gleichmäßige und so schwache Biegung des Oberrandes der Orbita hat, wohl nur am neuen Schädel aus der weiteren Umgebung von Kapstadt eine Parallele.
Hier möchte ich nun meinen Standpunkt, bezüglich so manchen anatomischen Factums an dem neuen Schädel klar umreißen: Solange uns die Architekturen z. B. der menschlichen Orbita und ihre dynamischen Bedingtheiten derart. unbekannt sind, muß ich als Anatom jede Erörterung darüber, ob die vorliegende Form der Orbita einen Neandertaler als ehemaligen Besitzer annehmen läßt oder nicht, als absurd bezeichnen. Wie viele Orbitaforuien zeigt uns Homo sapiens allein heute! Will man etwa die paar bis heute bekannten Orbitaformen des Homo neandertalensis als „die“ Neandertalerfor-men erklären? Wenn man sich als älterer Anatom und An-thropologe die ganz ungeheure Variabilität aller menschlichen Organe und Strukturen vor Augen hält, dann gehört Mut dazu, dennoch den Homo sapiens vom Homo neanderta-lensis scheiden zu wollen. Beide Formen gehörten ganz verschiedenen Zeiträumen an. Solange wir nicht Äpfel von Birnbäumen pflücken, solange kann ein Neandertaler gesun-der Konstitution unmöglich ein Sapienskind sein oder ein Sapiens ein Neandertalerkind!
Hier steht doch wohl der Wegweiser für jeden Naturforscher! Die erfolgreichsten unserer Vordermänner haben den Weg gebahnt. An uns ist es, ihn fortzubauen. Dem will diese Arbeit alleine die-nen! Irren ist menschlich, den großen Problemen auswei-chen, ist feige.
Die Vordermänner haben uns eine bestimmte Koppelung von Schädeleigentümlichkeiten als Unterscheidungs-merkmale zwischen Homo sapiens KING und seinem Nachfolger Homo sapiens LINNÉ errungen. Der Gedanke, daß nahezu jede Formeigenschaft typisch für den einen oder den anderen Homo sei, kann gar nicht erwogen werden. Suchen wir gemeinsam den s c h e i d e n d e n Komplex der Formgestaltung klarzulegen!
Der mesoconche Orbitalindex unseres Fundstückes steht mit 80,6 dem von La Chapelle nahe genug und der hypsiconche der Frau von Gibraltar bleibt mit 91,1 hinter denjenigen moderner Eskimo zurück! Das sollte zur Vorsicht mahnen.
Unsere Erörterung über den Torus und seine Kau-druckzuleitung wollen wir mit einer kurzen Betrachtung des erhaltenen Gebißanteils abschließen. Wenn im para-bolischen Zahnbogen. dem die drei vorderen Zähne fehlen, die Kauflächenlänge nur 710 mm beträgt, ist dies ein bescheidenes Maß für einen Neandertaler. Freilich ist eine sehr starke Abkauung festgestellt, wie so oft bei Nean-dertalerzähnen. So dürfte die Kauflächenlänge einst gewiß etwas länger gewesen sein. Für uns ist von Belang, daß dieses Gebiß einst keinen stärkeren Ausbau des Gesichtspfeiler und des Torus erzwingen mußte, als sie der Frauenschädel von Rhünda vorführt 4).
Eine sehr schmale sphenoide Schädelform weist die Norma verticalis nach. Ergänzt man den fehlenden linken Torus supraorbitalis symmetrisch. so beträgt die ganze Toruslänge nur 80 mm. Setzt man dies Maß in Beziehung zur größten Hirnschädelbreite (140 mm), so bekommt, man die Verhältniszahl 57,1. Als ich die Norma verticalis eines der Kinderschädel von Krapina damit verglich, fand ich fast die gleiche Proportion. Aber der Schädel der Rhündaerin war durch sein Pubertätswachsturn weit länger geworden.
Im Aufblick auf den Torus tritt dessen ziemlich schwache Entfaltung, namentlich verglichen mit jener robuster männlichen Schädel hervor. Sie bleibt etwas unter der des Frauenschädels von Gibraltar. Der Torus des Jünglings von Le Moustier 5) erinnert wohl in der Stärke an den der Frau von Rhünda, doch fehlt ihm die kolbige laterale Anschwellung. Wir beachten, daß der Torus der Rhündaerin, trotz der geringen Stirnbreite, die schmale Orbita voll überdeckt, und daß die untere Knochenspangen-Umrahmung der Orbita, welche den jugalen mit dem nasalen
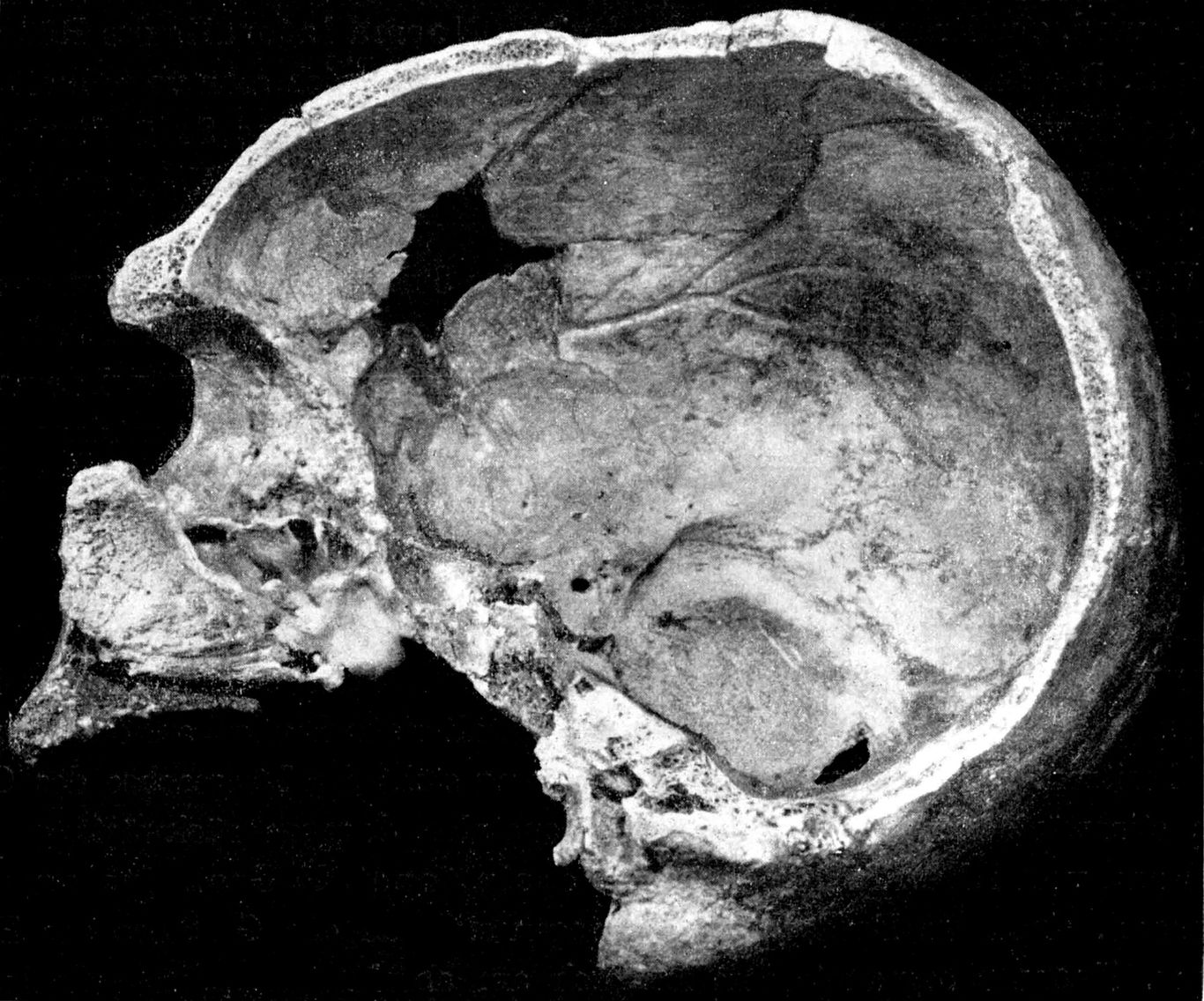
Abb. 8. Rhünda.Norma lateralis links. 1/2 natürlicher Größe,
Gesichtspfeiler in ihrer gegenseitigen Verbindung fixieren hilft, dank der so kräftigen Entfaltung des Processus frontosphenoides und des Jochbogens, dem Torus vollkommen parallel nach links zieht.
So ist uns klargeworden, daß der stumpfere Keilbeinwinkel der Neandertaler gegenüber dein Homo sapiens nicht allein den Hirnschädel platycephal gestaltete, sondern auch die so charakteristische Knochenbrille ihres Orbitaeinganges schuf. Die Ausgestaltung des Brillenoberrandes zum Torus supraorbitalis mit seinem nie fehlenden kräftigen Nasenbügel sowie der stabilere Apparat der drei Gesichtspfeiler und der sie seitwärts fixierende derbere Jochbogen, sie bilden die Kernstruktur des Gesichtsschädels, die nicht minder typisch für den Neandertaler ist.
Betrachten wir noch einmal KEITHs Schemazeichnungen (Abb. 4) und denken wir nun einmal den S a p i e n s s c h ä d e l aus Knetgummi konstruiert! Durch leichtes Anheben seiner Stirn und seines Hinterkopfes können wir ihn zu einem Neandertalerschädel umformen. Da wird uns verständlich, warum die Länge des Craniums der Neandertaler oberhalb des höchsten Mittelwertes heutiger Sapiensschädel gefunden wird. Wir begreifen nicht minder, daß der stumpfere Basiswinkel das Schädeldach flach macht.
Daß indessen die Schädeldachform nicht nur durch die Basisknickung bestimmt wird, studieren wir nun an der linken Norma lateralis des Rhündaer Schädels. Der Leser wird dann bemerken, daß es für mich sehr vorteilhaft war, daß das Unwetter vom 20. Juli 1956 den Einblick in dieses Schädelinnere ermöglicht hat. In der Schädelhöhle fällt zunächst die relative Höhe am Anfang des hinteren Abschnittes als fremdartig auf.
Wir werfen einen Blick auf den Verlauf des Sulcus transversus, der uns den alten Stand des Tentoriums cerebelli anzeigt. Wir denken uns die Höhe des einst vom sehr niederen Stirnlappen des Großhirns eingenommenen Schädelraums nun halbiert und ebenso die jenes Raumes, in dem einst der Hinterhauptslappen des Großhirns sich breitete. Durch beide Halbierungspunkte der Raumhöhe legen wir im Geiste die Großhirn-Längsachse. Sie steht von vorn nach hinten gesenkt!
Da gedenken wir der so bedeutsamen Entdeckung des Anatomen AUGUST FRORIEP vom Jahre 1897. FRORIEP zeigte uns, daß das Großhirn in zweierlei Lagerung angetroffen wird, die besonderen gestaltenden Einfluß auf das Schädeldach besitzen. Bei der frontopetalen Lagerung steht die Längsachse des Großhirns stirnwärts geneigt, bei der occipitopetalen dagegen rückwärts geneigt. Die Frau von Rhünda hat einst die occipitopetale Großhirnlagerung besessen. FRORIEPs Entdeckungen bezogen sich auf das Großhirn des Homo sapiens. FRORIEP begegnete ihm lediglich an, absolut gemessen, sehr langen Schädeln. Der Rhündaer Schädel ist solch ein sehr langer Schädel 4)!
Die Hauptmasse des Großhirns der Frau von Rhünda lag hinter dem Porus acusticus externus. Das Cavum cranii dieses Schädels konnte unter der stark geneigten Stirnbeinschuppe auf etwas Raum verzichten, denn der Gipfel des Großhirns lag hinter das vordere Drittel des Scheitelbeins verschoben. Deshalb mußten sich die hinteren zwei Drittel der Scheitelbeine etwas steiler stellen, als es bei weniger fliehender Stirn nötig gewesen wäre!
Beim Neandertaler ist, wie ich sehe, auch FRORIEPS zweiter Großhirnlagerungstyp schon gesehen worden, der frontopetale. KEITH hat ihn in Abb. 137 des zweiten Bandes seiner „Antiquity of Man“ vom Rhodesiaschädel vorgeführt, wenn er offenbar auch FRORIEPs Typen nicht gekannt hat.
Wichtig ist dieser Nachweis der Großhirntypen schon beim Neandertaler darum, weil er uns zeigt, daß auch am Schädeldach der Neandertaler bereits die Hirnlagerung formend mitgewirkt hat, wie heute bei Homo sapiens. Ohne Kenntnis dieser Dinge ist eine Beurteilung vieler Schädel gar nicht möglich!
Bekannt ist, daß an manchen Neandertalerschädeln Warzenfortsätze fehlen oder sehr schwach ausgebildet sind. Das gibt es ja heute bei Buschmännern, Hottentotten und bei Eskimos auch. Der bei Homo sapiens unterhalb des Porus acusticus allein vom Schläfenbein gebildete Spitzenteil des Processus mastoides ist beim Rhündaer Schädel 15 mm lang. Für drei Schläfenbeine aus Krapina teilte uns WEIDENREICH 1928 folgende Maße mit: 11, 13, 17 gegenüber der Ehringsdorferin mit 21 mm. Auch der Männerschädel von La Ferrassie sowie der La Quina-Schädel zeigen stattliche Warzenfortsätze.
Auf dem internationalen Kongreß „Hundert Jahre Neandertaler“ in Düsseldorf glaubte ein dem Fundstück ganz nahe sitzender Kollege, trotz der Stellung des Grundes der Fossa mandibularis auf etwa halber Gehörgangshöhe, trotz des durchaus typischen Torus supraorbitalis und der großen Schädellänge von 197 min, er habe einen mikrocephalen Sapiensschädel vor sich. Er suchte zu suggerieren, daß der Stirnlappen des Großhirns vorn zu liegen habe oder eben nicht entwickelt sei. Es stellte sich heraus, daß er von den FRORIEPschen Großhirntypen mit ihrer grundwichtigen Aussage auch nichts wußte.
Dabei war noch darauf hingewiesen, daß der Rhündaer Schädel keinen Sinus frontalis entwickelt hat. Sonst wäre die Länge dieses Fundstückes noch um einige Millimeter vergrößert!
Den Frauenschädel von Rhünda habe ich in Düsseldorf unter Heraushebung aller seiner Neandertaler-Platycephaliefolgen, sowohl im Hirn- wie im Gesichtsschädelbereich, an Hand von Lichtbildern erläutert. Zum Schluß habe ich die Eigenheiten seines Hinterhauptes aus dem Vorliegen der occipitopetalen Großhirnlagerung abgeleitet. Hingegen vermochte ich dem Vorkommen individueller Eigentümlichkeiten irn Rahmen der bislang ja relativ wenigen Neandertaler-Cranien keinen besonderen Wert beizumessen.
Da sich die erste Kritik des Rhündaer Schädels in Düsseldorf lediglich um Einzelheiten gekümmert hat, und von keiner Seite zu dem von mir so betonten Gesamtkomplex der Neandertalerschädel Stellung genommen worden ist, an dem ich das neue Fundstück maß, fasse ich nunmehr die Kernpunkte der Diagnostik zusammen:
1. Das platte Dach des Schädels von Homo neandertalensis KING entstammt vorwiegend einer stumpfwinkligeren Basisknickung als sie den erwachsenen Homo sapiens eignet.
2. Diese stumpfere Knickung drückt sich auch in der bedeutenderen Schädellänge aus. Sie liegt oberhalb des höchsten Mittelwertes heutiger Schädellängen.
3. Eine dritte Folge dieser Knickung ist die tiefere Lage des Grundes der Fossa mandibularis. Er steht in Höhe der Gehörgangmitte.
4. Sehr selten ist der Basiswinkel eines Neandertaler-Craniums direkt meßbar. Er konnte von SERA am Gibraltarschädel mit 140 o bestimmt werden. Da auch an diesem Cranium der Boden der Fossa mandibularis auf Gehörgangmitte stand, dürfte der Basiswinkel der Neandertaler im etwa 141° oscillieren, d. h. um den von WELCKER angegebenen Mittelwert der Basisknickung bei Neugeborenen des Homo sapiens.
5. Die Stärke des mandibularen Gelenkkopfes erzwang die Größe des typischen Basiswinkels der Neandertaler (KEITH).
Weitgehend gestützt auf das Schrifttum wurde weiter gezeigt, daß auch die Form des Gesichtsschädels der Neandertaler von der Basisknickung beherrscht ist. Die orbitale, vom Kieferdruck geschaffene, Knochenbrille - längst bekannt - ist mit ihrem derben Oberrande bei Homo sapiens ja überwiegend in die Stirnsteile des Knochens hineingenommen.
6. Der dünne Rand des fliehenden Stirnbeins des Neandertalerschädels wird erst durch energische Knochenauflagerung auf die Tabula externa ein leistungsfähiges Widerlager des Kieferdruckes, und durch Einbau feiner Knochenarkaden zum geeigneten Drucküberträger auf das flache Schädeldach. Durchlaufend verbindet der von einer Jochbeinnaht zur ande- ren ziehende Torus supra-orbitalis die beiden Jochbeinnähte des Stirnbeins. Er hat die Incisura frontalis lateralis ganz oder fast ganz durch seine Entfaltung zum Verschwinden gebracht, die Aufnahmestellen des höheren Druckes der Gesichts- pfeiler beiderseits verstärkt und oberhalb der Nasenwurzel den derben Nasenbügel des Torus heraustreten lassen.
7. Die Dachkonstruktion des Schädels der Neandertaler ist nicht allein vom stumpferen Keilbeinwinkel bestimmt. In ihr spielt außerdem der Lagerungstyp des Großhirns eine wichtige formende Rolle. FRORIEP zeigte sie uns einst an den Schädeln des Homo sapiens. KEITH belegte 1925 mit einer Abbildung das Bestehen des frontopetalen Hirntyps am Nean- dertaler-Cranium von Broken Hill, ich entdeckte am Rhündaer Schädel den occiipitopetalen. Die Existenz der beiden Großhirnlagerungstypen auch beim Neandertaler setzt der platycephalen Einstufung allein nach Stirn- und Lambdawinkel nunmehr Grenzen.
Die Diagnostik des Neandertaler-Cranium darf nur noch vom platycephalen Gesamtkomplex unter Berücksichtigung des Hirntypus in der Dachkonstruktion ausgehen. Denn es spielen in der erblich-fiaierten artlichen Besonderheit des Neandertalerschädels funktionelle Momente eine wesentliche Rolle und mahnen zur Behutsamkeit.
KINGs Namengebung hatte Homo neandertalensis zur Art erhoben. Heute können wir diese, wie mir scheint, dank so vieler erfolgreicher Vorarbeiten - bereits gegen Homo sapiens klar abgrenzen. Vielleicht sind wir dem Tage nicht mehr fern, an dem auch andere Glieder stammesgeschichtlicher Vergangenheit der Hominiden umgrenzbar werden.
Anmerkungen (für diese Fassung versetzt):
1) VIRCHOWs Arbeit war HUXLEY unbekannt geblieben.
2) 1) KEITH hat WELCKERs Arbeiten nicht benutzt.
3) Sie ist erforderlich, da wir ja nur einseitig kauen.
4) Über die bedeutende Kronenbreite des M 1 und M 2 vgl. S. 74.
5) Vgl. HAUSERS Photo auf Tafel 53 in F. MAURER: Der Mensch und seine Ahnen. Berlin, Ullstein 1928.
6) FRORIEPs finden sich auch in O Schulze: Topographische Anatomie, 4. Auflage, Abb. 72 u. 73.
Schrifttum.
ADLOFF, P., Das Gebiß des Menschen und der Anthropomorphen. Berlin 1908.
BOULE, M., L'Homme fossile de la Chapelle aux Saintes. L'Anthropologlie 19 (1908).
Gleicher Titel. In Annales de Paleóntologie 6 (1911).
CAPITAN et PEYRONIE, Station préhistorique de la Ferrassie. Rev. anthrop. 22 (1912).
FRAIPONT et LOHEST, La race humain de Neanderthal en Belgique. Arch. biol., Paris 7 (1887).
FRORIEP, A., Zur Kenntnis der Lagebeziehungen zwischen Großhirn und Schädeldach bei Menschen verschiedener Kopfform. Leipzig 1891.
DABELOW, A., Über Korrelationen in der phylogenetischen Entwicklung der Schädelform. II. Morph. Jb. 07 (1931).
GORNAJOVIC-KRAMBERGER, K., Der paläolithische Mensch von Krapina in Kroatien. Wiesbaden 1906.
GÖRKE, O., Beitrag zur funktionellen Gestaltung des Schädels bei den Anthropomorphen und Menschen durch Untersuchung mit Röntgenstrahlen. Arch. Anthrop. 1 (1904).
HRDLICKA, A., The Rhodesian Man. Amer. J. Physic. Anthrop. 9 (1926).
KING, W., The reputed fossil Man of the Neanderthal. Q. Journ. Sc. 1 (1861).
MARTIN, H., Recherches sur l´Evolution du Mousterien dans le gisement de la Quina 1-4.1926.
MARTIN, R., Lehrbuch der Anthropologie. Jena 1914.
MERKEL, F., Beitrag zur Kenntnis der postembryonalen Entwicklung des menschlichen Schädels. Festschrift für Henle 1882.
ROERIG, O ., Der Gesichtsteil des menschlichen Schädels. Arch. Entw. mech. Organ. 30 (1910).
SCHWALBE, G., Der Neandertalschädel. Bonner Jahrbücher. H. 106 (1901).
SERA, G., Per alcune sulla base del cranio. Arch. antrop., Florenz 41 (1922).
SERGI, S., Discovery of a cranium of Neanderthal typ near Rom. Zschr. Morph. Anthrop. 28 (1930).
SOBOTTA, J., Der Schädel von La Chapelle aux Saintes und die Mandibula des Homo heidelbergensis von Mauer. Zschr. Morph. Anthrop. 15 (1913).
TOLDT, C., Brauenwülste, Tori supraorbitales, und Brauenbogen, Arcus superciliares usw. und ihre morphologische Bedeutung. Mitt. Anthrop. Ges. Wien 1914.
WEIDENREICH, F., Die Sonderform des Menschenschädels in Anpassung an den aufrechten Gang. Zschr. Morph. Anthrop. 24 (1924).
-- Der Schädelfund von Weimar-Ehringsdorf. Jena 1928.
WEINERT, H., Der Schädel des eiszeitlichen Menschen von Le Moustier in seiner neuen Zusammensetzung. Berlin 1925.
-- Ursprung der Menschheit. Enke, Stuttgart 1932.
WELCKER, H., Untersuchungen über Wachstun und Bau des menschlichen Schädels. Leipzig 1862.
-- Über die Winkel der Schädelbasis. Verh. Ges. Dtseh. Naturf. Ärzte 1892.